科普驿站 第57期
主题:性——一个问题(中)
科目:生物
难度:B2
讲师:杜瑾鸿
对于细菌来说,质粒(plasmid)的交换和转位因子(transposable elements,TE)的跳动取代了性。前者是一种环状DNA,可被细菌吸收或释放(在细菌死亡后);后者则是小段的DNA或RNA序列,在几乎所有的细胞中都被发现,能在DNA(在这特指细菌)和质粒上移动,故又称跳跃基因,同时是一种诱变因子。质粒与转位因子的流动不但是菌株之间的,更是菌种之际的,这就是水平基因转移(horizontal gene transfer, HGT)。它正是细菌的抗药性的罪魁祸首。
对于原生动物来说,除去那些进行同配生殖的物种以外,性则意味着接合,即悄然靠近另外一个同类,溶解掉彼此的细胞壁,细胞质和细胞核相继混合。当它再度分裂为两个细胞时,就无法再区分彼此了。它的最初发现几乎瓦解了我们对性的认知。
不过这些还不是仅有的。人们曾考虑过梅塞尔松效应(Meselson Effect),它曾尝试预言克隆体通过独自积累突变,最终体现出更高的多样性。在贾第虫(Giardia)、甲螨(oribatid mite)、环柄菇科(Lepiotaceae)真菌、Darwinulid ostracods(一种介形虫)以及某些扁盘动物(Placozoa)等等这样曾经发生过从“有”到“无”的转变的候选对象身上,人们试图搜寻梅塞尔松效应,但并未发现。但巧合的是,人们发现了蛭形轮虫(Bdelloid rotifer)。
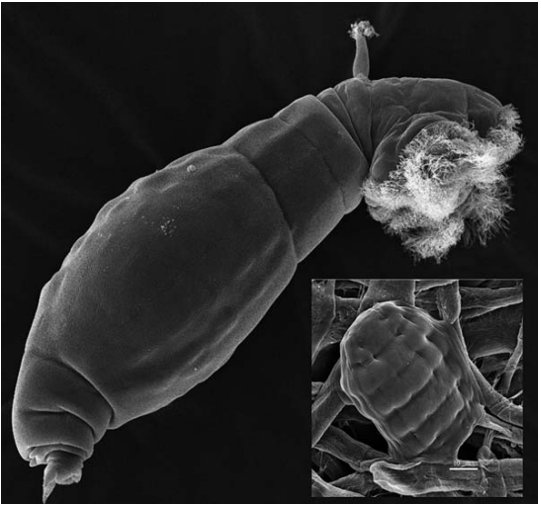
作为克隆体,它已经存在了几千万年,怪异的以至于J.M. Smith称之为“进化的丑行”(something of evolutionary scandal)。在Bdelloidea这个目中,至少有460个种。人们发现,蛭形轮虫的起源与几千万年前其二倍体祖先的全基因组复制事件有关,这导致蛭形轮虫变成了双二倍体(简并四倍体),并可能因此失去了性。
它与水熊虫十分相似,长度不到半毫米的身体装着复杂而完善的器官结构。它们能忍受大跨度的温度与酸碱度,全球性分布,几乎能在任何淡水环境中被找到,可以抵抗极端干旱与高能电离射线,对于拥有极强的DNA的重组和修复能力,可说是分布最广的微无脊椎动物之一。作为进化的怪胎,它引起了高度注意,其基因组、转录组(转录产物的总和,可以近似理解为mRNA)或是其他可能有核酸活跃的地方,都是潜在谜底。研究发现了多以转位因子的形式存在的外来(包括细菌、真菌、植物)序列;又暗示它存在大规模的水平基因转移;又发现蛭形轮虫可能在同属的种间存在(等位)基因共享(杂合化)……
我们似乎明白了什么。性本质上是一种基因交流(genetic mixing)。在一个无性种群中,自然选择会几乎以世系为单位清除不利个体,这将造成很多浪费。而一旦进行基因交流,其增益就很有可能立刻体现,这应该就是性,也即基因交流所提供的短期生存可能性。同时,从两种反面意义上,这种解释提供了更少的理论前提。
科学家还研究了似乎具有相同能力的另外一个物种——亚马逊帆鳍鲈。2018年2月,由七国学者组成的一个团队,在《自然》上联名发表研究,全面分析了另外这个无性的物种,并在它身上发现了远大于有性种群的多样性。
这种生活在墨西哥湾海域的鱼,需要相同地区其他有性近亲的精子来启动胚胎发育。但这个精子不会被吸纳,新个体仍然只是母亲的克隆体。研究估计,它的起源至少要追溯到10万年前(几十万代的克隆世系),这已远远超出了先前计算的预计灭绝时间(predicted extinction time, PET)。同时找出了它“失去了性”的原因:一次莫名的干扰把某次减数第一次分裂变成了(类)有丝分裂,创造了一个二倍体配子,从而开启了亚马逊帆鱼雌核生殖的进程。

人们同样在亚马逊帆鳍鲈的身上发现了水平基因转移,而转位因子可能发挥了关键作用,这或许正与它用别的动物的精子来触发自身胚胎发育的行为有关。然而当研究者考虑转位因子的存在时,重新计算的PET只多出了一丁点,根本无法解释10万年这个时间跨度。
为此,团队研究了另外一种基因交流的行为——种间杂交。实际上,大约10%的动物种群会有规律地与至少一个其他物种发生杂交。由性创造出来的多样性,是一个理由充足的起点,它赋予了转向克隆的突变体占领整个种群的能力,也带来了无限的危险。一旦转向克隆,杂交就是一种可能的生存策略。团队提出了冷冻杂合体基因组假说(frozen hybrid genome hypothesis),认为亚马逊帆鳍鲈正是一种冷冻杂合体(frozen hybrid)。研究认为:无性的失败不是因为克隆体相比于性有劣势,而是找不到能跟它杂交的有性物种。
这即说明,以复杂(而严酷的)环境条件作为前提,失去了性未必灭亡,但失去了基因共享必然灭亡。
参考引源:
(注:本参考文轩/书目名录中,有的是最初信息源,有的是二次或多次信息源(即引用了的部分不是该文献/书目的原创性结果),但一定能追溯到最初信息源。)
《英国通史》,主编:钱乘旦;作者:宋立宏,李家莉,张建辉等;卷六-篇三-章四.
《妇产科学》(第八版),主编:谢幸、苟文丽;人民卫生出版社;pp28.
《自私的基因》,[英]Richard Dawkins;
Smith J M. What use is sex?. Journal of Theoretical Biology, 1971, 30(2):319-335.
Gutekunst J, Andriantsoa R, Falckenhayn C, et al. Clonal genome evolution and rapid invasive spread of the marbled crayfish. Nature Ecology & Evolution, 2018, 2(3):567.
约翰·斯皮克曼, 张丽娜. 性的问题. 科学世界, 2017(3):136-137.
Smith J M. The evolution of sex. 1978.
Marek P E, Krejca J K, Shear W A. A new species of Illacme Cook & Loomis, 1928 from Sequoia National Park, California, with a world catalog of the Siphonorhinidae (Diplopoda, Siphonophorida)[J]. Zookeys, 2016, 626(626):1-43.
Kronauer D J C, Pierce N E, Keller L. Asexual reproduction in introduced and native populations of the ant Cerapachys biroi[J]. Molecular Ecology, 2012, 21(21):5221-5235.
《生殖生物学》,主编:窦肇华;人民卫生出版社.
Hörandl E. Meiosis and the Paradox of Sex in Nature. Intech, 2013.
Weismann, A. The Evolution Theory (Edward Arnold, London, 1904).
Kondrashov A S. Selection against harmful mutations in large sexual and asexual populations. Genetical Research, 1982, 40(3):325-332.
Kondrashov A S. Deleterious mutations and the evolution of sexual reproduction.. Nature, 1988, 336(6198):435-440.
Sohail M, Vakhrusheva O A, Sul J H, et al. Negative selection in humans and fruit flies involves synergistic epistasis. Science, 2016, 356(6337):539-542.
Paland S, Lynch M. Transitions to asexuality result in excess amino acid substitutions.[J]. Science, 2006, 311(5763):990-2.
Tucker A E, Ackerman M S, Eads B D, et al. Population-genomic insights into the evolutionary origin and fate of obligately asexual Daphnia pulex[J]. Proceedings of the National Academy of Sciences of the United States of America, 2013, 110(39):15740-5.
Goddard M R, Godfray H C J, Burt A. Sex increases the efficacy of natural selection in experimental yeast populations. Nature, 2005, 434(7033):636-40.
Auld S K, Tinkler S K, Tinsley M C. Sex as a strategy against rapidly evolving parasites. Proceedings of the Royal Society B Biological Sciences, 2016, 283(1845):20162226.
Smith J M.Contemplating life without sex. Nature, 1986, 324(6095):300-301.
Jokela J, Dybdahl M F, Lively C M. The maintenance of sex, clonal dynamics, and host-parasite coevolution in a mixed population of sexual and asexual snails. American Naturalist, 2009, 174 Suppl 1(S1):S43.
Schön, Isa, Martens, K. (Koen), Dijk, Peter van. Lost sex : the evolutionary biology of parthenogenesis. Springer, 2009.
Gladyshev E A, Meselson M, Arkhipova I R. Massive horizontal gene transfer in bdelloid rotifers. Science, 2008, 320(5880):1210-1213.
Gladyshev E, Meselson M. Extreme Resistance of Bdelloid Rotifers to Ionizing Radiation. Proceedings of the National Academy of Sciences of the United States of America, 2008, 105(13):5139-44.
Ricci C, Melone G, Santo N, et al. Morphological response of a bdelloid rotifer to desiccation.[J]. Journal of Morphology, 2003, 257(2):246.
Mark Welch D B, Mark Welch J L, Meselson M. Evidence for degenerate tetraploidy in bdelloid rotifers. Proc Natl Acad Sci U S A, 2008, 105(13):5145-5149.
Marotta R, Ricci C, Melone G. Morphological Changes Accompanying Anhydrobiosis in the Rotifer Macrotrachela quadricornifera (Rotifera, Bdelloidea). Journal of Morphology, 2008, 269(12):1481-1481.
Rodriguez F, Kenefick A, Arkhipova I. LTR-Retrotransposons from Bdelloid Rotifers Capture Additional ORFs Shared between Highly Diverse Retroelement Types. Viruses, 2017, 9(4):78.
Warren W C, Garcíapérez R, Xu S, et al. Clonal polymorphism and high heterozygosity in the celibate genome of the Amazon molly.. Nat Ecol Evol, 2018, 2(4).
Mallet J. Hybridization as an invasion of the genome.[J]. Trends in Ecology & Evolution, 2005, 20(5):229-237.
陈玉华. 端粒酶与肿瘤的研究进展. 湖北理工学院学报, 2010, 26(2):43-45.
李升伟. 端粒酶逆转衰老过程. 世界科学, 2011(1):49-50.
孔令平, 汪华侨. 端粒和端粒酶与衰老、癌症的潜在关系——2009年诺贝尔生理学或医学奖简介. 自然杂志, 2009, 31(6):327-331.
Archibald,J.The band of biologists who redrew the tree of life, Nature 560, 26-27 (2018).
闵嘉德. 合作造就的性. 科学世界, 2017(8):140-141.
辛迪·梅斯顿, 戴维·巴斯, CindyMeston,等. 女人的性爱动机. 2014.
Figure origins:
第1面:网络
第3面:Schön, Isa, Martens, K. (Koen), Dijk, Peter van. Lost sex : the evolutionary biology of parthenogenesis. Springer, 2009.
第4面上:维基百科
第4面下:网络
第5面上、下:网络

![[s-3]](https://www.bokeyuan.net/pic/image/emoji/cas/3.png)