科普驿站 第三十七期
主题:核酸的剪刀——CRISPR-Cas系统
科目:生物
难度:B2
讲师:杜瑾鸿
靶向RNA的CRISPR/Cas系统?
其实你早就应该意识到,CRISPR/Cas9不仅是一项基因编辑技术,在得知可以切割RNA的CRISPR/Cas系统之后就更是如此了,例如,2014年就发现,酿脓链球菌CRISPR/Cas9系统(即CRISPR/SpCas9,当前最广泛使用的CRISPR/Cas9)可以对单链RNA进行切割,但它作为RNA剪切的工具缺点太明显而且难以改进,(另外一个版本的CRISPR/Cas9——CRISPR/FnCas9也因为几乎相同的缺点而被抛弃。)所以并没有成为主要的RNA编辑工具,人们后来探索出了CRISPR/Cas13这一RNA编辑工具。CRISPR/Cas13属于VI-A型,主要有Cas13a,Cas13b两种,两者的作用机制基本相同,可以剪切单链RNA。此外,对CRISPR/Cas13d的研究与发掘也在进行。细菌的CRISPR/Cas13系统被细菌用来对抗RNA噬菌体。如今CRISPR/Cas13正在成为RNA编辑工具方面的领头雁,我们拭目以待。此外,由于CRISPR/Cas9的各种各样的不足,比如它的脱靶效应,系统效率,还有编辑能力在不同物种间的较大波动,乃至潜在的致癌性(见上文),还有其他种种,除了上述这些改造方法之外,人们还寻求将其他的CRISPR/Cas系统改造成更加高效的核酸编辑技术。
请你自行上网了解RNA靶向的CRISPR/Cas编辑技术。
其他的CRISPR/cas:为什么选择CRISPR/cas9?
细心的读者在阅读前文时就已经注意到,其实CRISPR/Cas系统是有多种的。目前已知的CRISPR/Cas系统可分成I,II,III,IV,V,VI六型,其中I,III,IV三型(type)归为1类(class),II,V,VI三型归为2类。读者不妨猜测,划分为1类和2类的标志是什么呢?
其实,划分1类和2类CRISPR/Cas的标准就是看它的效应物的结构是不是特别简单。我们把执行对抗二次入侵功能的蛋白质叫作效应物。第2类CRISPR/Cas系统的效应物无一不是一个蛋白质就能单枪匹马完成所有的剪切任务的。而第1类CRISPR/Cas则是由多个蛋白组成复合体来执行切割功能的。例如,第I型(属第1类)CRISPR/Cas的效应物就是多个Cas蛋白组成CRISPR相关病毒防御复合物(CRISPR associated complex for antivirus defense,Cascade)。(说起来也挺有意思的,cascade就有“大簇的下垂物”的意思。这就好像苏格兰场的犯罪数据库也要拼成Home Office Large Major Enquiry System(HOLMES)一样。)而III型亦类属第1类,它则是形成Cmr(III-B)或Csm(III-A)复合物来发挥效应的,这样好几个蛋白质组成的复合体庞大而复杂。可以说,因为Cas9只有一个蛋白质,且当时人们只清晰地了解前三型CRISPR/Cas系统,所以CRISPR/Cas9率先获得了青睐并被改造成核酸编辑技术。人们已经发现了6型22种主要的CRISPR/Cas系统类型,它们的基因组成、运行机制、效应物的分子结构乃至进化路径等的简介见于两篇文章:
SnapShot:Class 1 CRISPR-Cas Systems(doi:10.1016/j.cell.2017.02.018)
SnapShot:Class 2 CRISPR-Cas Systems(doi:10.1016/j.cell.2016.12.038)
各种CRISPR/Cas系统的标志性基因分别是:
I型CRISPR/Cas系统:Cas3;
可进一步分为I-A(Cas8a,Cas5)、I-B(Cas8b)、I-C(Cas8c)、I-D(Cas10d)、I-E(Cse1,Cse2)、I-F(Csy1, Csy2,Csy3)和I-U(GSU0054(cas5与cas6的融合基因))。
II型CRISPR/Cas系统:Cas9;
可进一步分为II-A(Csn2)、II-B(Cas4)和II-C(只有Cas9,Cas1,Cas2,不存在Csn2和Cas4)。
III型CRISPR/Cas系统:Cas10;
可进一步分为III-A(Csm2)、III-B(Cmr5)、III-C(Cas10或Csx11)和III-D(Csx10)。
IV型CRISPR/Cas系统:Csf1;
V型CRISPR/Cas系统:Cas12a(Cpf1)或Cas12b(C2c1)或Cas12c(C2c3);
可进一步分为V-A(Cas12a)、V-B(Cas12b)、V-C(Cas12c)和V-U(TnpB-like)
VI型CRISPR/Cas系统:Cas13a(C2c2)或Cas13b或Cas13c;
可进一步分为VI-A(Cas13a1(C2c2))、VI-B(Cas13b)和VI-C(cas13a2)。
就算是相同种类的Cas蛋白,在不同种类的细菌中也是不同的。所以诸如Cas9、Cas12a、Cas12b、Cas13a等这样的蛋白,其实还能更加细分出不同的种类,也就是说,每一种Cas蛋白都“坐拥一片土地”。那么科学家何以将Cas13分为Cas13a,Cas13b,Cas13c,Cas13d,又何以将Cas13a分为PspCas13a、LwaCas13a等等呢?你能根据这些CRISPR/Cas系统的基因组成来说明“标志性基因”这一概念的确切含义吗?
新的CRISPR/Cas基因组编辑工具
1. CRISPR/Cas12a
CRISPR/Cas12a系统属于V-A型,首先在新凶手弗朗西斯菌(Francisella novicida)和普雷沃菌(Prevotella)中被发现与Cas9同时存在。它的基因座结构如图所示。2015年,它被改造成基因编辑工具;2016年,它的效应物的晶体结构被完全解析。

引自SnapShot:Class 2 CRISPR-Cas Systems(doi:10.1016/j.cell.2016.12.038)
Cas12a与Cas9有以下不同:
Cas12a(也即Cpf1)比Cas9要小一些,它没有HNH结构域,而有一个NUC结构域,结构域的不同导致它切割产生的是5个核苷酸突出的黏性末端而Cas9产生的是平端。(原本因为只发现了RuvC结构域,导致科学家猜测Cas12a是形成二聚体来发挥功能的,直到它的晶体结构被解析。)
天然CRISPR/Cas12a系统的效应物中就只有一个RNA,Cas9需结合tracrRNA-crRNA——两个RNA。
Cas12a参与了pre-crRNA的剪切(换言之即crRNA的产生)。
CRISPR/Cas12a的效应物的crRNA的核苷酸数比Cas9进行编辑所需要的sgRNA小得多。(前者是42-44个核糖核苷酸,而后者需要>100个。)所以gRNA更加容易设计。
Cas9所识别的PAM序列通常富含G,而Cas12a所识别的PAM序列则富含T。
CRISPR/Cas12a在PAM序列下游的第23个碱基处,以及其互补链上的下游的第18个碱基处切割;Cas9在PAM上游的第3个碱基处切割。
Cas9在成功识别将近一半碱基后,就开始与目标DNA结合,而Cas12a会将几乎所有的碱基比对一遍,确认是否匹配,然后才与目标DNA结合。而且如果Cas12a发现不匹配的地方,会立刻脱离。(这点是因为它的R-loop结构。)
crRNA-Cas12a复合物在识别PAM序列时,能与PAM序列形成R-loop结构,这是一种螺旋结构,在形成完整的R-loop以后,就会进行剪切。
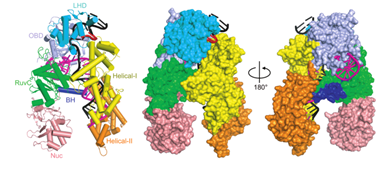
引自Gao P , Yang H , Rajashankar K R , et al. Type V CRISPR-Cas Cpf1 endonuclease employs a unique mechanism for crRNA-mediated target DNA recognition[J]. Cell Research, 2016.
因为以上种种原因,CRISPR/Cas12a系统的脱靶率将比Cas9低得多,并且因为它与Cas9的诸多不同,它将能与CRISPR/Cas9互补,从而扩大CRISPR/Cas系统的工具箱,使基于CRISPR/Cas系统的基因编辑技术具有更强的普适性和编辑能力。下图展示了这种工具的剪切过程。
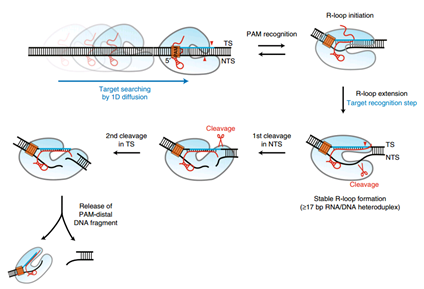
引自Yongmoon J , Hee C Y , Yunsu J , et al. Direct observation of DNA target searching and cleavage by CRISPR-Cas12a[J]. Nature Communications, 2018, 9(1):2777-.
你认为除此之外,Cas12a还有什么优点?
2.CRISPR/Cas12b
今年2月,Nature Communizations发表了张峰团队的Engineering of CRISPR-Cas12b for human genome editing,报道了第三种可以编辑人类基因组的CRISPR/Cas系统即CRISPR/Cas12b。Cas12b蛋白家族有27个成员。作者在论文中使用了外村尚芽孢杆菌(Bacillus hisashii)Cas12b(BhCas12b)来对人类基因组进行编辑。不过,在体温下,野生BhCas12b“优先切割非靶DNA链而不是形成双链断裂”,导致它切割效率降低。因此作者构建了优化突变体,并在体外进行人T细胞基因编辑实验,结果显示Cas12b比SpCas9(即CRISPR/Cas9基因技术使用的Cas9蛋白)体现出更高的特异性(脱靶率更低)。此外,因为Cas12b (也即C2c1)比Cas9和Cas12a更小(BhCas12b只有1108个氨基酸,LbCas12a(Lachnospiraceae bacterium ND2006 Cas12a)有1228个,而SpCas9含有1368个氨基酸),更容易通过病毒载体实现递送。
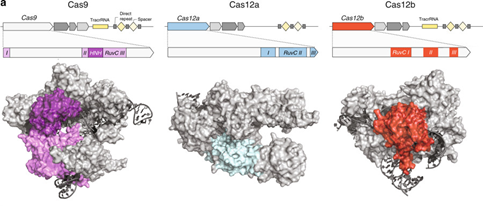
引自F Zhang.et al.Engineering of CRISPR-Cas12b for human genomeediting[J]. Nature Communications.2019
3.CRISPR/Cas3
你知道为什么第1类CRISPR/Cas会因效应物过于庞大而几乎没有用于基因编辑吗?
第1类中I型在所有发现存在CRISPR/Cas系统的细菌中分布最为广泛(约占了一半)。正如我们先前所介绍的,I型CRISPR/Cas系统的效应物是一个复合物Cascade。转录生成的pre-crRNA先结合在Cas6上,再被剪切成crRNA。crRNA-Cas6随后募集其他4种的Cas蛋白。
在大肠杆菌(I-E型)中,生成的crRNA-Cas6会募集1个Cse蛋白、2个Cse2蛋白、6个Cse7蛋白和1个Cas5蛋白。6个Cse7蛋白起到了支架作用。此后,对靶标DNA的切割则是由一个有解旋酶、核酸酶双重功能的蛋白质Cas3来执行的。(所以,我们也可以称I型CRISPR/Cas系统为CRISPR/Cas3系统。)要募集Cas3来执行它的功能,crRNA必须与靶标DNA完全互补配对,并形成R-loop结构,这样一来,Cas3才会被募集到Cascade。
事实上,crRNA与靶DNA之间的配对区竟可以长达32个碱基,I型CRISPR/Cas系统对于增加特异性、降低脱靶效应具有巨大的潜力;此外,I型CRISPR/Cas系统与第2类CRISPR/Cas系统不同,I型的切割靶点有很多个,分布于PAM上游的广大区域。也就是说,它可以在识别并定位到靶序列后,将PAM上游的所有DNA全部切碎成小段,这样的远程切割功能,是第2类CRISPR/Cas不具有的。这些特征让CRISPR/Cas3系统在降低脱靶率、提高特异性和切割效率方面具有很大潜力。但碍于Cascade复合体过于庞大,难以递送进入细胞核,所以它的可应用性一直无法在真核细胞中得到证明。
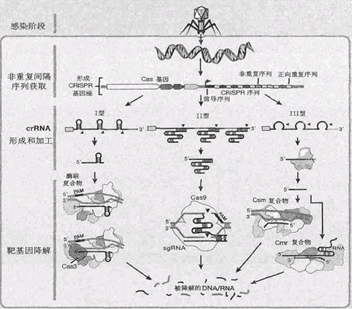
引自宁静, 李明煜, 鲁凤民. CRISPR/Cas9介导的基因组编辑技术[J]. 生物学通报, 2016(4):1-5.
今年4月,两个小组联合在Molecular Cell 上发表了文章Introducing a Spectrum of Long-Range Genomic Deletions in Human Embryonic Stem Cells Using Type I CRISPR-Cas,报道了使用噬热单胞菌(Thermobifida fusca)的CRISPR/Cas3进行基因编辑。不过,科研人员使用了比较复杂的办法来将庞大的Cascade复合物送入细胞核,并在切割结束后在PAM上游检测到了成千上万种不同的基因大片段缺失。“该系统可以更高效地清除人类基因组中的目标长链DNA,还可以帮助研究人员检测基因组中的非编码基因元件。”CRISPR/Cas3一次最多可以清除10万个脱氧核苷酸长度的DNA,充分地显示出了它的大片段删除能力。
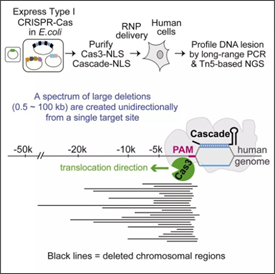
引自Adam E.Dolan,Zhonggang Hou,et al.Introducing a Spectrum of Long-Range Genomic Deletions in Human Embryonic Stem Cells Using Type I CRISPR-Cas[J]Molecular Cell.(2019),74,936-950
为什么科学家认为CRISPR/Cas3系统具有Cas9和Cas12所不具有的长片段删除能力?
4.CRISPR/AID
中科院上海生命科学院/上海交大医学院健康科学研究所的常兴团队利用CRISPR/dCas9与活化诱导胞嘧啶核苷脱氨酶(activation-induced cytidine deaminase,AID)结合,而AID可以制造碱基置换突变:单独作用时,可以将胞嘧啶或鸟嘌呤转换成其他碱基;与尿嘧啶DNA糖基化酶抑制剂联合使用可以定向地将胞嘧啶转化为胸腺嘧啶,而dCas9则起到定位作用。这一系统又一次扩展了CRISPR/Cas的功能范围。这一结果2016年10月10日发表在Nature Methods上,标题是Targeted AID-mediated mutagenesis (TAM) enables efficient genomic diversification in mammalian cells。
小结与展望
如今已经证明,CRISPR/Cas的最原始意义虽然是细菌的免疫系统,但在某些细菌中CRISPR/Cas演化出了其他功能。例如在条件致病菌铜绿假单胞菌(Pseudononas aeruginosa)中,已有实验证明了CRISPR/Cas系统能够调控生物膜的形成;再如粘细菌黄色粘球菌(Myxococcus xanthus),当它的一个CRISPR位点被转座子插入突变后,细胞不能聚集并发育形成子实体,这表明CRISPR/Cas系统与粘细菌的发育有关。还有研究发现Cas1参与DNA修复,如2011年发现纯化的双链断裂修复酶RecBCD 中包括 Cas1,且敲除Cas1后的大肠杆菌对一些抗生素更加敏感了,说明细菌和我们一样,懂得挖掘CRISPR/Cas的潜能或改造它。现在,CRISPR的变体和改造层出不穷,例如SunTag——这是一套分子挂钩,其能够将多个拷贝的生物活性分子挂到可用来靶向一些基因或其他的分子的蛋白质支架上,利用SunTag能增加CRISPR的变化。对它的改造使得CRISPR的功能和应用范围不断拓宽。
CRISPR/Cas系统如今用于核酸编辑、表达调控、突变追踪、基因扫描、应对抗药性细菌,甚至分子显微镜,从而帮助人类寻找新的基因、构建肿瘤,动物,神经疾病和传染病等研究模型、活细胞基因/蛋白显像、遗传病基因治疗、抵抗病毒、分子检测、农作物改良、物种保护、灭绝物种复活,乃至提高产能效率等,在各方面获得了广泛的应用。
CRISPR/Cas技术自身也在取得更多的改良发展。而且客观上,因为它兴起的时间较晚近,所以我国科学家在CRISPR/Cas这一领域在国际上取得了举足轻重的地位。总的来说,在CRISPR/Cas这一方面的研究和应用目前还处于起步状态,而且CRISPR/Cas还有很多缺陷和不足需要改进。本文完成时尚属2019年年中,发表时,恐怕已经因大量的新进展而显得有些过时。可以相信在未来,我们可以最大限度地发挥CRISPR/Cas的潜能,为人类创造更多财富。
参考资料:
Haft D H , Selengut J , Mongodin E F , et al. A Guild of 45 CRISPR-Associated (Cas) Protein Families and Multiple CRISPR/Cas Subtypes Exist in Prokaryotic Genomes[J]. PLoS Computational Biology, 2005, 1(6):e60.
Makarova K S , Haft D H , Barrangou R , et al. Evolution and classification of the CRISPR–Cas systems[J]. Nature Reviews Microbiology, 2011, 9(6):467-477.
在2005年的研究中,科学家首次为Cas蛋白做了分类,将45种蛋白归入鉴定出的10个亚型:核心蛋白(core proteins)、Ecoli亚型、Ypest亚型、Nmeni亚型、Dvulg亚型、Tneap亚型、Hmari亚型、Apern亚型、Mtube亚型、RAMP组件一共十个家族中。
Ecoli亚型(cse)来源于一个大肠杆菌品系:Escherichia coli K12-MG1655。
Ypest亚型(csy)来源于多个鼠疫耶尔森菌耶尔森菌(Yersinia pestis)品系。
Nmeni亚型(csn)来源于一个脑膜炎奈瑟氏球菌品系:Neisseria meningitidis Serogroup A Z2491。
Dvulg亚型(csd)来源于希尔登堡脱硫弧菌(Desulfovibrio vulgaris Hildenborough)。
Tneap亚型(cst)来源于一个新阿波罗栖热袍菌品系:Thermotoga neapolitana DSM4359。
Hmari亚型(csh)来源于一个死海盐盒菌品系:Haloarcula marismortui ATCC 43049。
Apern亚型(csa)来源于一个敏捷气热菌品系:Aeropyrum pernix K1。
Mtube亚型(csm)来源于两个结核分枝杆菌品系:Mycobacterium tuberculosis CDC1551和H37Rv。
RAMP组件(RAMP module)亚型(cmr)是一个特殊的Cas蛋白家族。
虽然在2011年进行了重新分类,但是很多蛋白根据该分类方法所赋予的名称代号却被沿用了下来,但也有很多蛋白的名字被更改。如Cas9曾被称作Csn1或Csx12。更多详细的资料可参见TIGRFAMs(
【本文为耀星会的原创作品,未经允许,禁止盗用、转载、篡改文章,否则耀星会将追究版权责任。】

![[s-70]](https://www.bokeyuan.net/pic/image/emoji/cas/70.png)











