科普驿站 第64期
主题:性——一个问题(下)
科目:生物
难度:B2
讲师:杜瑾鸿
(接第57期)另外一个方面,是避开基因交流的个体会被清除。考虑一种极具诱惑性的行为。冒着莫大的风险与另外一个个体杂交,与维持我身上的运转良好的基因组合,哪个更好一些呢?在克隆与性的纷争中,也正存在这样的逻辑。如果这样的作弊行为毫无限制,自诩清高就会成为进化主流,种群中不再产生新的个体,结果不是跟克隆体一样就是比克隆体更惨。生命进化出很多防止这种行为的机构装置。交配时强烈的快感,还有复杂的性反应机制,以及衰老都是其例。前两个显而易见,衰老何从谈起呢?
有句老话说:“肾上腺素决定出不出手,多巴胺决定天长地久,5-羟色胺决定谁先开口,端粒酶决定谁会先走。”2010年,科学家成功地逆转了小鼠的衰老,这可能是个令人振奋的好消息,但也可能意味着一场灾难。
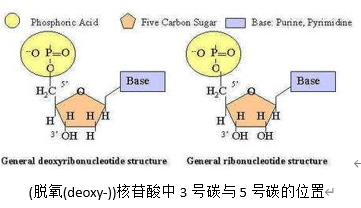
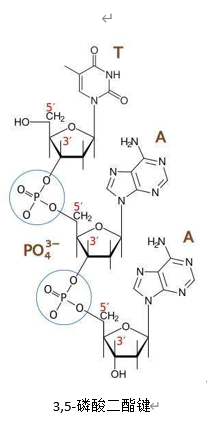
DNA的双链是反向互补的。相邻核苷酸之间通过3,5-磷酸二酯键连接,这个名称意在表示它是五碳糖中,5号碳上的磷酸基(上的一个羟基)与3号碳上的(醇)羟基结合形成的。所以链端会分别出现裸露的磷酸基和羟基,分别称5’端和3’端。聚合酶只能单向复制,且需要一个往往几个到几十个碱基长的核酸作引物,引物由DNA聚合酶α合成。对于旧双链中的其中一条,复制起点是旧链的3’端(新链的5’端),聚合酶ε以之为模板,畅通无阻地合成一条5’→3’的新链;另一条旧链的起点是5’端,聚合酶无法合成。
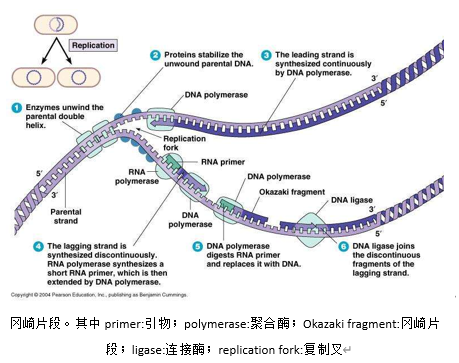
细胞解决这个问题的方法堪称巧妙,它先在该链起点前面的某处置一小段RNA作引物,聚合酶δ合成一小段DNA,再一直合成到复制起点;再放引物,再合成DNA;再放,再合成……最后,就出现了许多被RNA隔开的小段DNA(冈崎片段)。然后这些RNA被清除,聚合酶再以冈崎片段为引物合成剩余的链。但链仍不完整。空缺在于两端。而端粒就是用以修复它的。
2009年,诺贝尔生理学或医学奖被授予Elizabeth Blackturn、Carol Creider和Jack Szostak,以表彰他们在这方面的贡献。端粒出现在染色体的末端并不携带遗传信息,表现为某个固定重复的序列(如人类是TTAGGG),其最重要的结合物或相关蛋白是人端粒重复序列结合因子(telomeric repeat factor,TRF),分为TRF1和TRF2。前者对端粒长度有减损作用,且可抑制端粒酶的行为;TRF2则可以防止染色体末端相互融合,对维持端粒的正常结构必不可少。每复制一次,端粒就舍己为人,丢失一段。这样,端粒逐渐耗尽,细胞将渐趋衰老,因此它被看作是生命的时钟。而端粒酶则可以修复端粒。
人端粒酶广义上来说是一种核酸-蛋白复合物,由互补于端粒DNA的RNA亚基(human telomere RNA,hTR)和端粒酶相关蛋白(telomere associated protein,TEP)(如hTERT,hsp90,hEst1A和hEst1B等)组成。这样一来你可能也快猜到这些部分是干什么的了。hTERT是关键结构和主要调控亚单位,由1132个残基组成,能逆转录hTR合成端粒并添加到染色体端。自1985年在嗜热四膜虫(Tetrahymena thermophila)中找到了端粒酶后,人们发现端粒酶活性在癌细胞等不死性细胞和受精卵(胚胎细胞)中存在,而在大多数正常体细胞中无法检测到端粒酶活性。故对于正常细胞来说,存在一个时限(称Hayflick极限)使之达到后失去复制能力,从而逐渐衰老(和死亡),而只有接合才能让时钟重置。这样就能清除不进行基因交流的个体。
这样的障碍并不是仅有的。实际上,在雌雄同体生物身上,无论是渐渗杂交还是自体受精,最终结果都只能是一个世系在若干代以后变得完全同质。蚯蚓是雌雄同体生物,雌雄生殖器十分接近,但无论如何弯曲都无法自体受精。有些植物的花朵虽然雌雄蕊皆有,但要么是化学或物理上的障碍,要么是成熟时机不同阻碍了自体受精。
阻碍机制的存在,再次把我们导向了由它引起的灾难性后果。基因共享的手段还不存在时,是什么让生命得以延续?也许可以另辟蹊径式地认为,共享一开始就无处不在,那时甚至没有个体可言。只有当性的稳固机制确立起来以后,个体才获得了立足之地。让我们回到几十亿年前,岩浆味奶油彗星佳酿。20世纪生物学最伟大的思想家与创新者Carl Woese在20多年前曾提出过,生命可能起源于一种没有边界的“超有机体”(superorganism),那时,所有的生命都共享基因和新陈代谢。而性或者其他方式,则其实是在边界与隔离出现之后,仍继续进行基因交流的手段。事实上,生命从未放弃共享,我们看到不应该是进化树,而是“进化网”。这个观点也被David Quammen支持,他在2018年8月出版的著作《交横的树:生命新史》(The Tangled Tree: A Radical New History of Life)中,给出了它最具说服力的一批证据。
再来说性。性(尤其是两性)是作为一种最容易实现的基因共享的方式进化出来的。从水熊虫、蛭形轮虫两例能够看到,或许找到合适的杂交适配体能够导致的成功将是巨大的,但这样的几率太小了。可以想象,边界、多细胞生物的出现,都是对基因共享的打击。为此,通常只有不同的个体相互接触,才能使具有现实意义的基因共享成为可能。这最终使我们重新回到了Weismann,作为增加多样性的方式是一种表象,多样性的唯一途径是基因共享,性则是基因共享的最重要方式。其次,为了减小损耗,性就十有八九要与繁殖联系在一起,甚至成为繁殖的方式。在多细胞动物(尤其是结构复杂的高等动物)身上,性也作为一种决定新个体基因组的方式存在,因为任何对成年个体中个别细胞的改变,其效果往往都是微乎其微的。显然,性为什么是繁殖方式,和性有什么好处是两个问题。“性不必与繁殖联系在一起”,但这样一来,就不会再称作性了。
再次,它是在多细胞生物,尤其是在更高等的动物中,作为一种个体选择基因共享的对象的方式进化出来的。雌雄异体、体内受精可能只是早期的预兆,而配偶选择(行为)的意图则非常明显。如果你依然觉得过于抽象,还可以举个更加具体的例子。位于人类6号染色体的主要组织相容性复合体(Major histocompatibility complex, MHC)含有负责人体免疫功能的基因,具有广泛的多态性(polymorphism)。MHC基因相异的男女所生的孩子相对更加健康。研究发现,男女双方MHC基因的异同与相互吸引存在关联。MHC相异的双方将会对对方体味产生好感,情欲更加旺盛,婚姻更加稳定(不易出轨)。反之,这些指标全都降低。这个基因被广泛地研究,因为它被怀疑间接地驱动了多样性。
综上来说,性不愧是大自然鬼斧神工的伟大设计。基于此,性变成了生物界中最纷繁的构造之一。(2016年发现的一种“四根千足虫”,有4根阴茎(确切地说,生殖肢)。)但同时,它需要一定的支撑条件,如足够大的种群和足够丰富的基因库,这正是物种越珍惜灭绝越快的原因之一。性的意义已被我们找到,那么性的起源呢?遗憾的是,不像可以在实验室里煮酒论生命起源那样,我们找不到什么明显的迹象以及证据。不过,性的神秘面纱已渐被揭开,革命因故才要开始。
参考引源:
(注:本参考文轩/书目名录中,有的是最初信息源,有的是二次或多次信息源(即引用了的部分不是该文献/书目的原创性结果),但一定能追溯到最初信息源。)
《英国通史》,主编:钱乘旦;作者:宋立宏,李家莉,张建辉等;卷六-篇三-章四.
《妇产科学》(第八版),主编:谢幸、苟文丽;人民卫生出版社;pp28.
《自私的基因》,[英]Richard Dawkins;
Smith J M. What use is sex?. Journal of Theoretical Biology, 1971, 30(2):319-335.
Gutekunst J, Andriantsoa R, Falckenhayn C, et al. Clonal genome evolution and rapid invasive spread of the marbled crayfish. Nature Ecology & Evolution, 2018, 2(3):567.
约翰·斯皮克曼, 张丽娜. 性的问题. 科学世界, 2017(3):136-137.
Smith J M. The evolution of sex. 1978.
Marek P E, Krejca J K, Shear W A. A new species of Illacme Cook & Loomis, 1928 from Sequoia National Park, California, with a world catalog of the Siphonorhinidae (Diplopoda, Siphonophorida)[J]. Zookeys, 2016, 626(626):1-43.
Kronauer D J C, Pierce N E, Keller L. Asexual reproduction in introduced and native populations of the ant Cerapachys biroi[J]. Molecular Ecology, 2012, 21(21):5221-5235.
《生殖生物学》,主编:窦肇华;人民卫生出版社.
Hörandl E. Meiosis and the Paradox of Sex in Nature. Intech, 2013.
Weismann, A. The Evolution Theory (Edward Arnold, London, 1904).
Kondrashov A S. Selection against harmful mutations in large sexual and asexual populations. Genetical Research, 1982, 40(3):325-332.
Kondrashov A S. Deleterious mutations and the evolution of sexual reproduction.. Nature, 1988, 336(6198):435-440.
Sohail M, Vakhrusheva O A, Sul J H, et al. Negative selection in humans and fruit flies involves synergistic epistasis. Science, 2016, 356(6337):539-542.
Paland S, Lynch M. Transitions to asexuality result in excess amino acid substitutions.[J]. Science, 2006, 311(5763):990-2.
Tucker A E, Ackerman M S, Eads B D, et al. Population-genomic insights into the evolutionary origin and fate of obligately asexual Daphnia pulex[J]. Proceedings of the National Academy of Sciences of the United States of America, 2013, 110(39):15740-5.
Goddard M R, Godfray H C J, Burt A. Sex increases the efficacy of natural selection in experimental yeast populations. Nature, 2005, 434(7033):636-40.
Auld S K, Tinkler S K, Tinsley M C. Sex as a strategy against rapidly evolving parasites. Proceedings of the Royal Society B Biological Sciences, 2016, 283(1845):20162226.
Smith J M.Contemplating life without sex. Nature, 1986, 324(6095):300-301.
Jokela J, Dybdahl M F, Lively C M. The maintenance of sex, clonal dynamics, and host-parasite coevolution in a mixed population of sexual and asexual snails. American Naturalist, 2009, 174 Suppl 1(S1):S43.
Schön, Isa, Martens, K. (Koen), Dijk, Peter van. Lost sex : the evolutionary biology of parthenogenesis. Springer, 2009.
Gladyshev E A, Meselson M, Arkhipova I R. Massive horizontal gene transfer in bdelloid rotifers. Science, 2008, 320(5880):1210-1213.
Gladyshev E, Meselson M. Extreme Resistance of Bdelloid Rotifers to Ionizing Radiation. Proceedings of the National Academy of Sciences of the United States of America, 2008, 105(13):5139-44.
Ricci C, Melone G, Santo N, et al. Morphological response of a bdelloid rotifer to desiccation.[J]. Journal of Morphology, 2003, 257(2):246.
Mark Welch D B, Mark Welch J L, Meselson M. Evidence for degenerate tetraploidy in bdelloid rotifers. Proc Natl Acad Sci U S A, 2008, 105(13):5145-5149.
Marotta R, Ricci C, Melone G. Morphological Changes Accompanying Anhydrobiosis in the Rotifer Macrotrachela quadricornifera (Rotifera, Bdelloidea). Journal of Morphology, 2008, 269(12):1481-1481.
Rodriguez F, Kenefick A, Arkhipova I. LTR-Retrotransposons from Bdelloid Rotifers Capture Additional ORFs Shared between Highly Diverse Retroelement Types. Viruses, 2017, 9(4):78.
Warren W C, Garcíapérez R, Xu S, et al. Clonal polymorphism and high heterozygosity in the celibate genome of the Amazon molly.. Nat Ecol Evol, 2018, 2(4).
Mallet J. Hybridization as an invasion of the genome.[J]. Trends in Ecology & Evolution, 2005, 20(5):229-237.
陈玉华. 端粒酶与肿瘤的研究进展. 湖北理工学院学报, 2010, 26(2):43-45.
李升伟. 端粒酶逆转衰老过程. 世界科学, 2011(1):49-50.
孔令平, 汪华侨. 端粒和端粒酶与衰老、癌症的潜在关系——2009年诺贝尔生理学或医学奖简介. 自然杂志, 2009, 31(6):327-331.
Archibald,J.The band of biologists who redrew the tree of life, Nature 560, 26-27 (2018).
闵嘉德. 合作造就的性. 科学世界, 2017(8):140-141.
辛迪·梅斯顿, 戴维·巴斯, CindyMeston,等. 女人的性爱动机. 2014.
Figure origins:
第1面:网络
第3面:Schön, Isa, Martens, K. (Koen), Dijk, Peter van. Lost sex : the evolutionary biology of parthenogenesis. Springer, 2009.
第4面上:维基百科
第4面下:网络
第5面上、下:网络













不如想一想目前人类文明之中。
尤其是那些较为发达的国家。
越发达的国家的人为什么越来越就不想?
生育?
找到根源所在了之后要如何去解决?
哪怕理论上能够做到。
现实中那些统治者们
估计也不会做。